Чем отличается ядро прокариот от ядра эукариот. В чем отличие эукариот от прокариот? Сравнение прокариот и эукариот
Все живые организмы могут быть распределены в одну из двух групп (прокариоты или эукариоты) в зависимости от основной структуры их клеток. Прокариоты - живые организмы, состоящие из клеток, которые не имеют клеточного ядра и мембранных органелл. Эукариоты - живые организмы, которых содержат ядро, а также мембранные органеллы.
Клетка является фундаментальной составляющей нашего современного определения жизни и живых существ. Клетки рассматриваются в качестве основных строительных блоков жизни и используются в определении того, что значит быть «живым».
Давайте взглянем на одно определение жизни: «Живые существа - это химические организации, состоящие из клеток и способные размножаться» (Китон, 1986). Это определение базируется на двух теориях - клеточной теории и теории биогенеза. впервые была предложена в конце 1830-х годов немецкими учеными Маттиасом Якобом Шлейденом и Теодором Шванном. Они утверждали, что все живые существа состоят из клеток. Теория биогенеза, предложенная Рудольфом Вирховым в 1858 году, утверждает, что все живые клетки возникают из существующих (живых) клеток и не могут появиться спонтанно из неживой материи.
Компоненты клеток заключены в мембрану, которая служит барьером между внешним миром и внутренними составляющими клетки. Клеточная мембрана - избирательный барьер, это означает, что он пропускает некоторые химические вещества, поддерживающие равновесие, необходимое для жизнедеятельности клеток.
Клеточная мембрана регулирует перемещение химических веществ из клетки в клетку следующими способами:
- диффузия (тенденция молекул вещества к минимизации концентрации, то есть перемещение молекул из области с более высокой концентрацией по направлению к области с более низкой до момента выравнивания концентрации);
- осмос (движение молекул растворителя через частично проницаемую мембрану для того, чтобы уравнять концентрацию растворенного вещества, которое не в состоянии двигаться через мембрану);
- селективный транспорт (при помощи мембранных каналов и насосов).
Прокариоты - организмы, состоящие из клеток, которые не имеют клеточного ядра или любых мембранных органелл. Это означает, что генетический материал ДНК у прокариот не связан в ядре. Кроме того, ДНК прокариот менее структурирована, чем у эукариот. В прокариотах ДНК одноконтурная. ДНК эукариот организована в хромосомы. Большинство прокариот состоят только из одной клетки (одноклеточные), но есть несколько и многоклеточных. Ученые разделяют прокариот на две группы: и .
Типичная клетка прокариота включает:
- плазматическую (клеточную) мембрану;
- цитоплазму;
- рибосомы;
- жгутики и пили;
- нуклеоид;
- плазмиды;
Эукариоты

Эукариоты - живые организмы, клетки которых содержат ядро и мембранные органеллы. Генетический материал у эукариот находится в ядре, а ДНК организована в хромосомы. Эукариотические организмы могут быть одноклеточными и многоклеточными. являются эукариотами. Также эукариоты включают растения, грибы и простейших.
Типичная клетка эукариота включает:
- ядрышко;
1. У прокариот отсутствуют мембраны, ограничивающие органеллы бактериальной клетки(ядро. митохондрии, рибосомы) от цитоплазмы. Из мембран имеется только цитоплазматическая мембрана.
2. Ядро прокариот (нуклеоид) фибриальной структуры, ядерная оболочка отсутствует.
3. У прокариот отсутствуют митохондрии, хлоропласты, КГ. ЭПС.
4. Окислительно-восстановительные фрагменты локализованы в мезосомах (производных цитоплазматической мембраны)
5. У прокариот отсутствует митоз, размножаются путем бинарного деления.
6. Прокариоты имеют гаплоидный геном.
7. Отсутствует клеточный центр
8. Внутриклеточные перемещения цитоплазмы и амебоидное движение для прокариот нетипичны.
Специфические черты М/О
1. Малые размеры, масса, объем и относительная простота строения.
2. Чрезвычайно высокие темпы размножения
3. Большое разнообразие способов получения энергии и путём обмена веществ, широкий спектр конечных продуктов метаболизма.
4. Способность к биодеструкции практически всех естественных и искусственных веществ.
5. Чрезвычайно высокая степень адоптации как результат высоких темпов изменчивости.
6. Массовая популяция и повсеместное распространение.
6. Структура и функции поверхностных образований бактериальной клетки. Капсула. Методы выявления.
Бактериальная клетка окружена внешней оболочкой (рис. 3.2), которая состоит из капсулы, капсулоподобной оболочки и клеточной стенки. От их состава зависит способность клетки воспринимать анилиновые красители (тинкториальные свойства). Капсулы в зависимости от степени выраженности подразделяют на микро- и макрокапсулы. Первые обнаруживаются только при электронно-микроскопическом исследовании в виде микрофибрилл из мукополисаха-ридов, которые тесно прилегают к клеточной стенке. Макрокапсулы представляют собой выраженный слизистый слой, снаружи покрыва ющий клеточную стенку. Он состоит из полисахаридов и редко из полипептидов (например, у сибиреязвенных бактерий). Как правило макрокапсулу образуют немногие виды патогенных бактерий (пнев мококки и др.) при неблагоприятных условиях среды, например в организме животных или человека. Однако у некоторых видов (клебсиеллы пневмонии) макрокапсула обнаруживается постоянно.
Капсулоподобная оболочка - липидо-полисахаридное образование, сравнительно непрочно связанное с поверхностью клетки, вследствие чего в отличие от капсулы может выделяться в окружающую среду.
Капсула или капсулоподобная оболочка может быть покрыта экзополисахаридами, которые образуются из углеводов окружающей среды под действием бактериальных ферментов. При этом глюканы и леваны обеспечивают прилипание бактерий к разным поверхностям, часто гладким.
Капсула несет различные функции:
1. Защитная, предохраняя клетку от неблагоприятных условий среды обитания,
2. адгезивная, способствуя «прилипанию» к поверхносш (рецепюрам) клетки хозяина.
3. Часто патогенные и антигенные свойства. Непатогенные бактерии также могут образовывать макрокапсулу, выполняющую, по-видимому, только защитную функцию.
7. Структура и функции клеточной стенки грамположительных и грамотрицательных бактерий. Формы бактерий с дефектами клеточной стенки.
Клеточная стенка (КС)- биогетерополимер сложною химического состава, покрывающий всю поверхность прокариотической клетки.
Основа клеточной стенки- пептидогликан, обеспечивающий ригидность и эластичность КС. Структура пептидогликана паралелльные полисахаридные(гликановые) цепи состоящие из чередующихся звеньев [\"-ацетил1 лнжозамина и N-ацетилмурамовой к-ты С каждым остатком N-ацетидмурамовой к-ты ковалентно связан трипептид
Различии между Грам+ и Грам- бактериями.
| Группа | 1"рамм -*-_^____________________ | Грамм - |
| Окраска по Граму | фиолетовые | розовые |
| Толщина КС | 20-60 нм | 10-20 нм |
| % содержание липидов | 1,6% | 22,6% |
| Структура пептидогликана | Пептиды пептидогликанов связаны через пептидильный мостик из 5 , остатков глицина | Ацетилмурамовые к-ты каждой гликановой цепи связаны через два однотипных тетрапептида |
| % содержание пептидогликана | 40-90% Многослойный | 5-10% Однослойный |
| Наличие тейхоевых кислот | Имеются | Отсутствуют |
| Особеность выделения ферментов | Ферменты выделяются непоследственно в окружающую среду | Ферменты выделяются в периплазматическое пространство, находящееся между КС и ЦМ |
| Представители | Все болезнетворные кокки, кроме гонокка и менингококка, бациллы, иклострпдии | Энтеробактерии, вибрионы, трепонемы |
Функции КС:
1. Придает клетке определенную форму.
2. Защищает её от воздействия окружающей среды
3. Несет на поверхности разнообразные рецепторы, к которым прикрепляются некоторые фаги, колишины и химические соединения.
4. Через КС в клетку поступают питательные вещества и выделяются продукты обмена
5. Сдерживает высокое внутриклеточное осмотическое давление.
КС грам- бакт. представлена трехслойной внешней мембраной (пептидогликан + липополисахарид + липопротеиды). Некоторые белки (норины), пронизывая внешнююмембрану,образуют поры, через которую проходят гидрофильные в-ва с низкой молекулярной массой.
Пептидогликан- мишень действия некоторых антибиотиков (пенициллина) и ферментов (лизоцима). Пенициллин нарушает образование тетрапепдидных связей, лизоцим разрушает гликозидные связи между мурамовой к-той и ацетилглюкозамином.
При действии пенициллина на на растущую бак. культуру образуются безоболочечные формы бактерий:
1 Пртопласты- полностью лишены КС.
2.Сферопласты- частично лишены КС
И протопласты, и сферопласты подвергаются плазмолизу в изотонической среде, н в пшерюнической среде проявляютслабую метаболическую активность, ! утрачивают способность к размножению.
3.L- формы- полностью или частично лишены КС, сохраняют способность к размножению.
а) стабильные- способны к реверсии в исходный вид.
б) нестабильный-не способны к реверсии
8. Цитоппазматические структуры бактерий, функции, методы выявления. Кислотоустойчивые микробы. Метод окраски.
Жгутики . На поверхности ряда бактериальных клеток располагаются жгутики (рис. 3.5). В их состав входит белок флагелин, который по своей структуре относится к сократимым белкам типа Миозина. Жгутики прикрепляются к базальному телу, состоящему из системы нескольких дисков, вмонтированных в цитоплазматическую мембрану и КС. Количество и расположение жгутиков у разных бактерий неодинаково.
Монотрихи имеют на одном из полюсов клетки только один жгутик,
лофотрихи - пучок жгутиков,
амфитрихи - жгутики расположены на обоих полюсах клетки,
перитрихов - По всей ее поверхности.
Жгутики обладают антигенными свойствами.
Пили - тонкие полые нити белковой природы длиной 0,3-10 мкм, толщиной 10 нм, покрываю щие поверхность бактериальных клеток. В отличие от жгутиков не выполняют локомоторную функцию. По своему функциональному назначению подразделяются на несколько типов.
Пили 1 общего типа обусловливают прикрепление или адгезию бактерий к определенным клеткам организма хозяина. Их количество велико - от нескольких сотен до нескольких тысяч на одну бактериальную клетку. Адгезия является первоначальной стадией любого инфекционного процесса.
Пили 2 типа (синоним: конъюгативные, или половые, пили - sex pili) участвуют в конъюгации бактерий обеспечивающей перенос части генетического материала от донорной клетки к реци-пиентной. Они имеются только у бактерий-доноров в ограниченном количестве (1-4 на клетку).
Цитоплазматическая мембрана (ЦМ) является жизненно необходимым структурным компонентом бактериальной клетки. Она ограничивает протопласт, располагаясь непосредственно под клеточной стенкой. ЦМ в химическом отношении представляет собой липопротеин, состоящий из 15-30% липидов и 50-70% протеинов. Кроме того, в ней содержится около 2-5% углеводов и незначительное количество РНК. В состав мембранных липидов входят главным образом нейтральные липиды и фосфолипиды. У некоторых бактерий встречаются гликолипиды, а у микоплазм - стеролы.
Липидный состав мембран непостоянен в качественном и количественном отношении. У одного и того же вида бактерий он изменяется в зависимости от условий ее культивирования на питательной среде и возраста культуры. Разные виды бактерий отличаются друг от друга по липидному составу своих мембран.
Мембранные белки разделяются на структурные и функциональные. К последним относятся ферменты, участвующие в биосинтезе разных компонентов КС, который происходит на поверхности ЦМ, а также окислительно-восстановительные ферменты, пермеазы и др.
ЦМ является сложно организованной структурой, состоящей из трех слоев, которые выявляются при электронно-микроскопическом исследовании. Двойной фосфолипидный слой пронизан глобулинами, которые обеспечивают транспорт веществ в бактериальную клетку.
ЦМ выполняют жизненно важные функции, нарушение которых приводит бактериальную клетку к гибели. К ним относится прежде всего регуляция поступления в клетку метаболитов и ионов, участие
в метаболизме, репликации ДНК, а у ряда бактерий в спорообразовании и т.д.
Мезосомы являются производными ЦМ. Они имеют неодинаковое строение у разных бактерий, располагаясь в разных частях клетки либо в виде концентрических мембран, либо пузырьков, трубочек, либо в форме петли, характерной в основном для грамотрица-тельных бактерий. Мезосомы связаны с нуклеоидом. Они участвуют в делении клетки и спорообразовании.
Цитоплазма у прокариот, так же как и у эукариот, представляет собой сложную коллоидную систему, состоящую из воды (около 75%), минеральных соединений, белков, РНК и ДНК, которые входят в состав органелл нуклеоида, рибосом, мезосом, включений.
Нуклеоид является эквивалентом ядра эукариот, хотя отличается от него по своей структуре и химическому составу. Он лишен ядерной мембраны, не содержит хромосом, не делится митозом. В составе нуклеоида отсутствуют основные белки - гистоны. Исключение составляют только некоторые бактерии. В нем содержится двунитевая молекула ДНК, а также небольшое количество РНК и белков. Молекула ДНК с молекулярной массой (2-3) х 10 9 представляет собой замкнутую кольцевую структуру, в которой закодирована вся наследственная информация клетки, т.е. геном клетки. По аналогии с хромосомами эукариот бактериальная ДНК часто обозначается как хромосома. При этом следует помнить, что она представлена в клетке в единственном числе, поскольку бактерии являются гаплоидными. Однако перед делением клетки число нуклеоидов удваивается, а во время деления увеличивается до 4 и более.
Наряду с нуклеоидом в цитоплазме могут находиться автономные кольцевые молекулы двунитевой ДНК с меньшей молекулярной массой, которые получили название плазмид. В них также закодирована наследственная информация. Однако она не является жизненно необходимой для бактериальной клетки.
Рибосомы у бактерий представляют собой рибонуклеопротеиновые частицы размером 20 нм, состоящие из двух субъединиц 30S и 50S. Перед началом синтеза белка происходит объединение этих субъединиц в одну - 70S. В отличие от клеток эукариотов рибосомы бактерий не объединены в эндоплазматическую сеть. Бактериальные рибосомы, являющиеся белоксинтезирующими системами клеток, могут стать «мишенью» для действия многих антибиотиков.
Включения являются продуктами метаболизма про- и эука-РИотических микроорганизмов, которые располагаются в их цитоплазме и используются в качестве запасных питательных веществ. К ним относятся включения гликогена, крахмала, серы, полифосфата (волютина) и др. У некоторых бактерий, например дифтерийной палочки, включения волютина имеют дифференциально-диагностическое значение. Они обладают способностью к метахромазии (окрашиваются в иной цвет, чем цвет красителя).
Важнейшая, основополагающая особенность эукариотических клеток связана с расположением генетического аппарата в клетке. Генетический аппарат всех эукариот находится в ядре и защищён ядерной оболочкой (по-гречески «эукариот» значит имеющий ядро). ДНК эукариот линейная (у прокариот ДНК кольцевая и находится в особой области клетки - нуклеоиде, который не отделён мембраной от остальной цитоплазмы). Она связана с белками-гистонами и другими белками хромосом, которых нет у бактерий.
В жизненном цикле эукариот обычно присутствуют две ядерные фазы (гаплофаза и диплофаза). Первая фаза характеризуется гаплоидным (одинарным) набором хромосом, далее, сливаясь, две гаплоидные клетки (или два ядра) образуют диплоидную клетку (ядро), содержащую двойной (диплоидный) набор хромосом. Иногда при следующем делении, а чаще спустя несколько делений клетка вновь становится гаплоидной. Такой жизненный цикл и в целом диплоидность для прокариот не характерны.
Третье, пожалуй, самое интересное отличие, - это наличие у эукариотических клеток особых органелл, имеющих свой генетический аппарат, размножающихся делением и окружённых мембраной. Эти органеллы - митохондрии и пластиды. По своему строению и жизнедеятельности они поразительно похожи на бактерий. Это обстоятельство натолкнуло современных учёных на мысль, что подобные организмы являются потомками бактерий, вступившими в симбиотические отношения с эукариотами. Прокариоты характеризуются малым количеством органелл, и ни одна из них не окружена двойной мембраной. В клетках прокариот нет эндоплазматического ретикулума, аппарата Гольджи, лизосом.
Ещё одно важное различие между прокариотами и эукариотами - наличие у эукариот эндоцитоза, в том числе у многих групп - фагоцитоза. Фагоцитозом (дословно «поедание клеткой») называют способность эукариотических клеток захватывать, заключая в мембранный пузырёк, и переваривать самые разные твёрдые частицы. Этот процесс обеспечивает в организме важную защитную функцию. Впервые он был открыт И. И. Мечниковым у морских звезд. Появление фагоцитоза у эукариот скорее всего связано со средними размерами (далее о размерных различиях написано подробнее). Размеры прокариотических клеток несоизмеримо меньше, и поэтому в процессе эволюционного развития эукариот у них возникла проблема снабжения организма большим количеством пищи. Как следствие среди эукариот появляются первые настоящие, подвижные хищники.
Большинство бактерий имеет клеточную стенку, отличную от эукариотической (далеко не все эукариоты имеют её). У прокариот это прочная структура, состоящая главным образом из муреина (у архей из псевдомуреина). Строение муреина таково, что каждая клетка окружена особым сетчатым мешком, являющимся одной огромной молекулой. Среди эукариот клеточную стенку имеют многие протисты, грибы и растения. У грибов она состоит из хитина и глюканов, у низших растений - из целлюлозы и гликопротеинов, диатомовые водоросли синтезируют клеточную стенку из кремниевых кислот, у высших растений она состоит из целлюлозы, гемицеллюлозы и пектина. Видимо, для более крупных эукариотических клеток стало невозможно создавать клеточную стенку из одной молекулы высокую по прочности. Это обстоятельство могло заставить эукариот использовать иной материал для клеточной стенки. Другое объяснение состоит в том, что общий предок эукариот в связи с переходом к хищничеству утратил клеточную стенку, а затем были утрачены и гены, отвечающие за синтез муреина. При возврате части эукариот к осмотрофному питанию клеточная стенка появилась вновь, но уже на другой биохимической основе.
Разнообразен и обмен веществ у бактерий. Вообще всего выделяют четыре типа питания, и среди бактерий встречаются все. Это фотоавтотрофные, фотогетеротрофные, хемоавтотрофные, хемогетеротрофные (фототрофные используют энергию солнечного света, хемотрофные используют химическую энергию).
План
Введение 2
Прокариотическая
клетка 4
Эукариотическая
клетка 6
Сопоставление
про- и эукариотической клеток 13
Отличия эукариот
от прокариот 14
Список использованной
литературы: 19
Введение
Клетка - элементарная единица строения и жизнедеятельности всех живых организмов (кроме вирусов, о которых нередко говорят как о неклеточных формах жизни), обладающая собственным обменом веществ, способная к самостоятельному существованию, самовоспроизведению и развитию. Все живые организмы либо, как многоклеточные животные, растения и грибы, состоят из множества клеток, либо, как многие простейшие и бактерии, являются одноклеточными организмами. Раздел биологии, занимающийся изучением строения и жизнедеятельности клеток, получил название цитологии. В последнее время принято также говорить о биологии клетки, или клеточной биологии.История открытия
Первым человеком, увидевшим клетки, был английский учёный Роберт Гук (известный нам благодаря закону Гука). В 1665 году, пытаясь понять, почему пробковое дерево так хорошо плавает, Гук стал рассматривать тонкие срезы пробки с помощью усовершенствованного им микроскопа. Он обнаружил, что пробка разделена на множество крошечных ячеек, напомнивших ему монастырские кельи, и он назвал эти ячейки клетками (по-английски cell означает «келья, ячейка, клетка»). В 1675 году итальянский врач М. Мальпиги, а в 1682 году - английский ботаник Н. Грю подтвердили клеточное строение растений. О клетке стали говорить как о «пузырьке, наполненном питательным соком». В 1674 году голландский мастер Антоний ван Левенгук (Anton van Leeuwenhoek, 1632-1723) с помощью микроскопа впервые увидел в капле воды «зверьков» - движущиеся живые организмы (инфузории, амёбы, бактерии). Также Левенгук впервые наблюдал животные клетки - эритроциты и сперматозоиды. Таким образом, уже к началу XVIII века учёные знали, что под большим увеличением растения имеют ячеистое строение, и видели некоторые организмы, которые позже получили название одноклеточных. В 1802-1808 годах французский исследователь Шарль-Франсуа Мирбель установил, что все растения состоят из тканей, образованных клетками. Ж. Б. Ламарк в 1809 году распространил идею Мирбеля о клеточном строении и на животные организмы. В 1825 году чешский учёный Я. Пуркине открыл ядро яйцеклетки птиц, а в 1839 ввёл термин «протоплазма». В 1831 году английский ботаник Р. Броун впервые описал ядро растительной клетки, а в 1833 году установил, что ядро является обязательным органоидом клетки растения. С тех пор главным в организации клеток считается не мембрана, а содержимое.
Клеточная теория строения организмов была сформирована в 1839 году немецким зоологом Т. Шванном и М. Шлейденом и включала в себя три положения. В 1858 году Рудольф Вирхов дополнил её ещё одним положением, однако в его идеях присутствовал ряд ошибок: так, он предполагал, что клетки слабо связаны друг с другом и существуют каждая «сама по себе». Лишь позднее удалось доказать целостность клеточной системы.
В 1878 году русским учёным И. Д. Чистяковым открыт митоз в растительных клетках; в 1878 году В. Флемминг и П. И. Перемежко обнаруживают митоз у животных. В 1882 году В. Флемминг наблюдает мейоз у животных клеток, а в 1888 году Э. Страсбургер - у растительных.
Строение клеток
Все клеточные формы жизни на Земле можно разделить на два надцарства на основании строения составляющих их клеток:
прокариоты (доядерные) - более простые по строению и возникли в процессе эволюции раньше;
эукариоты (ядерные) - более сложные, возникли позже. Клетки, составляющие тело человека, являются эукариотическими.
Несмотря на многообразие форм организация клеток всех живых организмов подчинена единым структурным принципам.
Содержимое клетки отделено от окружающей среды плазматической мембраной, или плазмалеммой. Внутри клетка заполнена цитоплазмой, в которой расположены различные органоиды и клеточные включения, а также генетический материал в виде молекулы ДНК. Каждый из органоидов клетки выполняет свою особую функцию, а в совокупности все они определяют жизнедеятельность клетки в целом.
Прокариотическая клетка
Прокариоты (от лат. pro - перед, до и греч. ?????? - ядро, орех) - организмы, не обладающие, в отличие от эукариот, оформленным клеточным ядром и другими внутренними мембранными органоидами (за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий). Единственная крупная кольцевая (у некоторых видов - линейная) двухцепочечная молекула ДНК, в которой содержится основная часть генетического материала клетки (так называемый нуклеоид) не образует комплекса с белками-гистонами (так называемого хроматина). К прокариотам относятся бактерии, в том числе цианобактерии (сине-зелёные водоросли), и археи. Потомками прокариотических клеток являются органеллы эукариотических клеток - митохондрии и пластиды. Основное содержимое клетки, заполняющее весь её объём, - вязкая зернистая цитоплазма.Прокариоты (лат. Procaryota, от др.-греч. ??? «перед» и?????? «ядро»), или доядерные - одноклеточные живые организмы, не обладающие (в отличие от эукариот) оформленным клеточным ядром и другими внутренними мембранными органоидами (за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий). Единственная крупная кольцевая (у некоторых видов - линейная) двухцепочечная молекула ДНК, в которой содержится основная часть генетического материала клетки (так называемый нуклеоид) не образует комплекса с белками-гистонами (так называемого хроматина). К прокариотам относятся бактерии, в том числе цианобактерии (сине-зелёные водоросли), и археи. Потомками прокариотических клеток являются органеллы эукариотических клеток - митохондрии и пластиды.
Прокариоты разделяют на два таксона в ранге домена (надцарства): Бактерии (Bacteria) и Археи (Archaea).
Для клеток прокариот характерно отсутствие ядерной оболочки, ДНК упакована без участия гистонов. Тип питания осмотрофный.
Генетический материал прокариот представлен одной молекулой ДНК, замкнутой в кольцо, имеется только один репликон. В клетках отсутствуют органоиды, имеющие мембранное строение. В геноме могут присутствовать мобильные генетические элементы, а у некоторых прокариот (например, вольбахия) их содержится необычно много. Изучение бактерий привело к открытию горизонтального переноса генов, который был описан в Японии в 1959 г. Это процесс широко распространен среди прокариот, а также у некоторых эукариот. Открытие горизонтального переноса генов у прокариот заставило по-другому взглянуть на эволюцию жизни. Ранее эволюционная теория базировалась на том, что виды не могут обмениваться наследственной информацией. Прокариоты могут обмениваться генами между собой непосредственно (конъюгация, трансформация) а также с помощью вирусов - бактериофагов (трансдукция).
Строение типичной клетки
прокариот: капсула, клеточная стенка,
плазмолемма, цитоплазма, рибосомы, плазмида,
пили, жгутик, нуклеоид.
Характерные особенности
-
Отсутствие четко оформленного ядра
Наличие жгутиков, плазмид и газовых вакуолей
Структуры, в которых происходит фотосинтез
Формы размножения - бесполый способ, имеется псевдосексуальный процесс, в результате которого происходит лишь обмен генетической информацией, без увеличения числа клеток.
Размер рибосомы - 70s(по коэф. седиментации различают и рибосомы др. типов, а также субчастицы и биополимеры, входящие в состав рибосом)
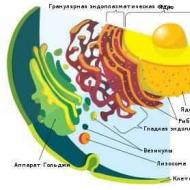
Эукариотическая клетка
Эукариоты (эвкариоты) (от греч. ?? - хорошо, полностью и?????? - ядро, орех) - организмы, обладающие, в отличие от прокариот, оформленным клеточным ядром, отграниченным от цитоплазмы ядерной оболочкой. Генетический материал заключён в нескольких линейных двухцепочных молекулах ДНК (в зависимости от вида организмов их число на ядро может колебаться от двух до нескольких сотен), прикреплённых изнутри к мембране клеточного ядра и образующих у подавляющего большинства (кроме динофлагеллят) комплекс с белками-гистонами, называемый хроматином. В клетках эукариот имеется система внутренних мембран, образующих, помимо ядра, ряд других органоидов (эндоплазматическая сеть, аппарат Гольджи и др.). Кроме того, у подавляющего большинства имеются постоянные внутриклеточные симбионты-прокариоты - митохондрии, а у водорослей и растений - также и пластиды.Эукариоты, или Ядерные (лат. Eukaryota от греч. ??- - хорошо и?????? - ядро) - домен (надцарство) живых организмов, клетки которых содержат ядра. Все организмы, кроме бактерий и архей, являются ядерными (вирусы и вироиды также не являются эукариотами, но не все биологи считают их живыми организмами).
Животные, растения, грибы, а также группы организмов под общим названием протисты - все являются эукариотическими организмами. Они могут быть одноклеточными и многоклеточными, но все имеют общий план строения клеток. Считается, что все эти столь несхожие организмы имеют общее происхождение, поэтому группа ядерных рассматривается как монофилетический таксон наивысшего ранга. Согласно наиболее распространённым гипотезам, эукариоты появились 1,5–2 млрд. лет назад. Важную роль в эволюции эукариот сыграл симбиогенез - симбиоз между эукариотической клеткой, видимо, уже имевшей ядро и способной к фагоцитозу, и проглоченными этой клеткой бактериями - предшественниками митохондрий и пластидов.
Эндомембранная система и её
компоненты
Строение эукариотической
клетки
Эукариотические клетки в среднем
намного крупнее прокариотических, разница
в объёме достигает тысяч раз. Клетки эукариот
включают около десятка видов различных
структур, известных как органоиды (или
органеллы, что, правда, несколько искажает
первоначальное значение этого термина),
из которых многие отделены от цитоплазмы
одной или несколькими мембранами (в прокариотических
клетках внутренние органоиды, окруженные
мембраной, встречаются редко). Ядро -
это часть клетки, окружённая у эукариот
двойной мембраной (двумя элементарными
мембранами) и содержащая генетический
материал: молекулы ДНК, «упакованные»
в хромосомы. Ядро обычно одно, но бывают
и многоядерные клетки.
Схематическое изображение
животной клетки. (При нажатии на
какое-либо из названий составных частей
клетки, будет осуществлён переход на
соответствующую статью.)
Поверхностный комплекс
животной клетки
Состоит из гликокаликса, плазмалеммы
и расположенного под ней кортикального
слоя цитоплазмы. Плазматическая мембрана
называется также плазмалеммой, наружной
клеточной мембраной. Это биологическая
мембрана, толщиной около 10 нанометров.
Обеспечивает в первую очередь разграничительную
функцию по отношению к внешней для клетки
среде. Кроме этого она выполняет транспортную
функцию. На сохранение целостности своей
мембраны клетка не тратит энергии: молекулы
удерживаются по тому же принципу, по которому
удерживаются вместе молекулы жира -
гидрофобным частям молекул термодинамически
выгоднее располагаться в непосредственной
близости друг к другу. Гликокаликс представляет
собой «заякоренные» в плазмалемме молекулы
олигосахаридов, полисахаридов, гликопротеинов
и гликолипидов. Гликокаликс выполняет
рецепторную и маркерную функции. Плазматическая
мембрана животных клеток в основном состоит
из фосфолипидов и липопротеидов со вкрапленными
в неё молекулами белков, в частности,
поверхностных антигенов и рецепторов.
В кортикальном (прилегающем к плазматической
мембране) слое цитоплазмы находятся специфические
элементы цитоскелета - упорядоченные
определённым образом актиновые микрофиламенты.
Основной и самой важной функцией кортикального
слоя (кортекса) являются псевдоподиальные
реакции: выбрасывание, прикрепление и
сокращение псевдоподий. При этом микрофиламенты
перестраиваются, удлиняются или укорачиваются.
От структуры цитоскелета кортикального
слоя зависит также форма клетки (например,
наличие микроворсинок).
Структура цитоплазмы
Жидкую составляющую цитоплазмы
также называют цитозолем. Под световым
микроскопом казалось, что клетка заполнена
чем-то вроде жидкой плазмы или золя, в
котором «плавают» ядро и другие органоиды.
На самом деле это не так. Внутреннее пространство
эукариотической клетки строго упорядочено.
Передвижение органоидов координируется
при помощи специализированных транспортных
систем, так называемых микротрубочек,
служащих внутриклеточными «дорогами»
и специальных белков динеинов и кинезинов,
играющих роль «двигателей». Отдельные
белковые молекулы также не диффундируют
свободно по всему внутриклеточному пространству,
а направляются в необходимые компартменты
при помощи специальных сигналов на их
поверхности, узнаваемых транспортными
системами клетки.
Эндоплазматический
ретикулум
В эукариотической клетке существует
система переходящих друг в друга мембранных
отсеков (трубок и цистерн), которая называется
эндоплазматическим ретикулумом (или
эндоплазматическая сеть, ЭПР или ЭПС).
Ту часть ЭПР, к мембранам которого прикреплены
рибосомы, относят к гранулярному (или
шероховатому) эндоплазматическому ретикулуму,
на его мембранах происходит синтез белков.
Те компартменты, на стенках которых нет
рибосом, относят к агранулярному (или
гладкому) ЭПР, принимающему участие в
синтезе липидов. Внутренние пространства
гладкого и гранулярного ЭПР не изолированы,
а переходят друг в друга и сообщаются
с просветом ядерной оболочки.
Аппарат Гольджи
Аппарат Гольджи представляет
собой стопку плоских мембранных цистерн,
несколько расширенных ближе к краям.
В цистернах аппарата Гольджи созревают
некоторые белки, синтезированные на мембранах
гранулярного ЭПР и предназначенные для
секреции или образования лизосом. Аппарат
Гольджи асимметричен - цистерны располагающиеся
ближе к ядру клетки (цис-Гольджи) содержат
наименее зрелые белки, к этим цистернам
непрерывно присоединяются мембранные
пузырьки - везикулы, отпочковывающиеся
от эндоплазматического ретикулума. По-видимому,
при помощи таких же пузырьков происходит
дальнейшее перемещение созревающих белков
от одной цистерны к другой. В конце концов
от противоположного конца органеллы
(транс-Гольджи) отпочковываются пузырьки,
содержащие полностью зрелые белки.
Ядро
Клеточное ядро содержит молекулы
ДНК, на которых записана генетическая
информация организма. В ядре происходит
репликация - удвоение молекул ДНК,
а также транскрипция - синтез
молекул РНК на матрице ДНК. В
ядре же синтезированные молекулы РНК
претерпевают некоторые модификации
(например, в процессе сплайсинга из
молекул матричной РНК исключаются незначащие,
бессмысленные участки), после чего выходят
в цитоплазму. Сборка рибосом также происходит
в ядре, в специальных образованиях, называемых
ядрышками. Компартмент для ядра - кариотека
- образован за счёт расширения и слияния
друг с другом цистерн эндоплазматической
сети таким образом, что у ядра образовались
двойные стенки за счёт окружающих его
узких компартментов ядерной оболочки.
Полость ядерной оболочки называется
люменом или перинуклеарным пространством.
Внутренняя поверхность ядерной оболочки
подстилается ядерной ламиной, жесткой
белковой структурой, образованной белками-ламинами,
к которой прикреплены нити хромосомной
ДНК. В некоторых местах внутренняя и внешняя
мембраны ядерной оболочки сливаются
и образуют так называемые ядерные поры,
через которые происходит материальный
обмен между ядром и цитоплазмой.
Лизосомы
Лизосома - небольшое
тельце, ограниченное от цитоплазмы одинарной
мембраной. В ней находятся литические
ферменты, способные расщепить все
биополимеры. Основная функция -
аутолиз - то есть расщепление отдельных
органоидов, участков цитоплазмы клетки.
Цитоскелет
К элементам цитоскелета
относят белковые фибриллярные структуры,
расположенные в цитоплазме клетки: микротрубочки,
актиновые и промежуточные филаменты.
Микротрубочки принимают участие в транспорте
органелл, входят в состав жгутиков, из
микротрубочек строится митотическое
веретено деления. Актиновые филаменты
необходимы для поддержания формы клетки,
псевдоподиальных реакций. Роль промежуточных
филаментов, по-видимому, также заключается
в поддержании структуры клетки. Белки
цитоскелета составляют несколько десятков
процентов от массы клеточного белка.
Центриоли
Центриоли представляют собой
цилиндрические белковые структуры, расположенные
вблизи ядра клеток животных (у растений
центриолей нет). Центриоль представляет
собой цилиндр, боковая поверхность
которого образована девятью наборами
микротрубочек. Количество микротрубочек
в наборе может колебаться для
разных организмов от 1 до 3.
Вокруг центриолей находится
так называемый центр организации
цитоскелета, район в котором группируются
минус концы микротрубочек клетки.
Перед делением клетка содержит
две центриоли, расположенные под
прямым углом друг к другу. В ходе
митоза они расходятся к разным концам
клетки, формируя полюса веретена деления.
После цитокинеза каждая дочерняя клетка
получает по одной центриоли, которая
удваивается к следующему делению. Удвоение
центриолей происходит не делением, а
путём синтеза новой структуры, перпендикулярной
существующей.
Центриоли, по-видимому, гомологичны
базальным телам жгутиков и ресничек.
Митохондрии
Митохондрии - особые органеллы
клетки, основной функцией которых
является синтез АТФ - универсального
носителя энергии. Дыхание (поглощение
кислорода и выделение углекислого
газа) происходит также за счёт энзиматических
систем митохондрий.
Внутренний просвет митохондрий,
называемый матриксом отграничен от
цитоплазмы двумя мембранами, наружной
и внутренней, между которыми располагается
межмембранное пространство. Внутренняя
мембрана митохондрии образует складки,
так называемые кристы. В матриксе содержатся
различные ферменты, принимающие участие
в дыхании и синтезе АТФ. Центральное значение
для синтеза АТФ имеет водородный потенциал
внутренней мембраны митохондрии.
Митохондрии имеют свой собственный
ДНК-геном и прокариотические рибосомы,
что безусловно указывает на симбиотическое
происхождение этих органелл. В ДНК митохондрий
закодированы совсем не все митохондриальные
белки, большая часть генов митохондриальных
белков находятся в ядерном геноме, а соответствующие
им продукты синтезируются в цитоплазме,
а затем транспортируются в митохондрии.
Геномы митохондрий отличаются по размерам:
например геном человеческих митохондрий
содержит всего 13 генов. Самое большое
число митохондриальных генов (97) из изученных
организмов имеет простейшее Reclinomonas americana.
Деление на царства
Существует несколько
вариантов деления надцарства эукариот
на царства. Первыми были выделены царства
растений и животных. Затем было выделено
царство грибов, которые из-за биохимических
особенностей, по мнению большинства биологов,
не могут быть причислены ни к одному из
этих царств. Также некоторые авторы выделяют
царства простейших, миксомицетов, хромистов.
Некоторые системы насчитывают до 20 царств.
По системе Томаса Кавалир-Смита все эукариоты
подразделяются на два монофилетических
таксона - Unikonta и Bikonta.
Деление эукариотических
клеток
Амитоз - прямое деление
клетки, происходит в соматических
клетках эукариот реже, чем митоз.
В большинстве случаев амитоз
наблюдается в клетках со сниженной
митотической активностью: это стареющие
или патологически измененные клетки,
часто обреченные на гибель (клетки
зародышевых оболочек млекопитающих,
опухолевые клетки и другие). При
амитозе морфологически сохраняется
интерфазное состояние ядра, хорошо видны
ядрышко и ядерная оболочка. Репликация
ДНК отсутствует. Спирализация хроматина
не происходит, хромосомы не выявляются.
Клетка сохраняет свойственную ей функциональную
активность, которая почти полностью исчезает
при митозе. Таково, например, деление
макронуклеусов многих инфузорий, где
без образования веретена происходит
сегрегация коротких фрагментов хромосом.
При амитозе делится только ядро, причём
без образования веретена деления, поэтому
наследственный материал распределяется
случайным образом. Отсутствие цитокинеза
приводит к образованию двуядерных клеток,
которые в дальнейшем не способны вступать
в нормальный митотический цикл. При повторных
амитозах могут образовываться многоядерные
клетки.
Митоз (от греч. ????? - нить)
- непрямое деление клетки, наиболее распространённый
способ репродукции эукариотических клеток,
один из фундаментальных процессов онтогенеза.
Митотическое деление обеспечивает рост
многоклеточных эукариот за счёт увеличения
популяции тканевых клеток. Биологическое
значение митоза заключается в строго
одинаковом распределении хромосом между
дочерними ядрами, что обеспечивает образование
генетически идентичных дочерних клеток
и сохраняет преемственность в ряду клеточных
поколений. Дробление оплодотворённого
яйца и рост большинства тканей у животных
также происходит путём митотических
делений. На основании морфологических
особенностей митоз условно подразделяется
на:
-
профазу,
прометафазу,
метафазу,
анафазу,
телофазу.
Мейоз (от греч. meiosis - уменьшение) или редукционное деление клетки - деление ядра эукариотической клетки с уменьшением числа хромосом в два раза. Происходит в два этапа (редукционный и эквационный этапы мейоза). Мейоз не следует смешивать с гаметогенезом - образованием специализированных половых клеток или гамет из недифференцированных стволовых. Уменьшение числа хромосом в результате мейоза в жизненном цикле ведёт к переходу от диплоидной фазы к гаплоидной. Восстановление плоидности (переход от гаплоидной фазы к диплоидной) происходит в результате полового процесса. В связи с тем, что в профазе первого, редукционного, этапа происходит попарное слияние (конъюгация) гомологичных хромосом, правильное протекание мейоза возможно только в диплоидных клетках или в чётных полиплоидах (тетра-, гексаплоидных и т. п. клетках). Мейоз может происходить и в нечётных полиплоидах (три-, пентаплоидных и т. п. клетках), но в них, из-за невозможности обеспечить попарное слияние хромосом в профазе I, расхождение хромосом происходит с нарушениями, которые ставят под угрозу жизнеспособность клетки или развивающегося из неё многоклеточного гаплоидного организма. Этот же механизм лежит в основе стерильности межвидовых гибридов. Определённые ограничения на конъюгацию хромосом накладывают и хромосомные мутации (масштабные делеции, дупликации, инверсии или транслокации).
Деление прокариотических клеток
Прокариотические клетки делятся надвое. Сначала клетка удлиняется, в ней образуется поперечная перегородка. На завершающем этапе дочерние клетки расходятся. Отличительной чертой деления прокариотических клеток является непосредственное участие реплицированной ДНК в процессе деления. Обычно прокариотические клетки делятся с образованием двух одинаковых по размеру дочерних клеток, поэтому этот процесс ещё иногда называют бинарным делением. В связи с тем, что в подавляющем большинстве случаев прокариотические клетки имеют клеточную стенку, бинарное деление сопровождается образованием септы - перегородки между дочерними клетками, которая затем расслаивается посередине. Процесс деления прокариотической клетки подробно изучен на примере Escherichia coli.
Сопоставление про- и эукариотической клеток
Наиболее важным отличием эукариот от прокариот долгое время считалось наличие оформленного ядра и мембранных органоидов. Однако к 1970-1980-м гг. стало ясно, что это лишь следствие более глубинных различий в организации цитоскелета. Некоторое время считалось, что цитоскелет свойственен только эукариотам, но в середине 1990-х гг. белки, гомологичные основным белкам цитоскелета эукариот, были обнаружены и у бактерий.Именно наличие специфическим образом устроенного цитоскелета позволяет эукариотам создать систему подвижных внутренних мембранных органоидов. Кроме того, цитоскелет позволяет осуществлять эндо- и экзоцитоз (как предполагается, именно благодаря эндоцитозу в эукариотных клетках появились внутриклеточные симбионты, в том числе митохондрии и пластиды). Другая важнейшая функция цитоскелета эукариот - обеспечение деления ядра (митоз и мейоз) и тела (цитотомия) эукариотной клетки (деление прокариотических клеткок организовано проще). Различия в строении цитоскелета объясняют и другие отличия про- и эукариот - например, постоянство и простоту форм прокариотических клеток и значительное разнообразие формы и способность к её изменению у эукариотических, а также относительно большие размеры последних. Так, размеры прокариотических клеток составляют в среднем 0,5-5 мкм, размеры эукариотических - в среднем от 10 до 50 мкм. Кроме того, только среди эукариот попадаются поистине гигантские клетки, такие как массивные яйцеклетки акул или страусов (в птичьем яйце весь желток - это одна огромная яйцеклетка), нейроны крупных млекопитающих, отростки которых, укрепленные цитоскелетом, могут достигать десятков сантиметров в длину.
Отличия эукариот от прокариот
Важнейшая, основополагающая особенность эукариотических клеток связана с расположением генетического аппарата в клетке. Генетический аппарат всех эукариот находится в ядре и защищён ядерной оболочкой (по-гречески «эукариот» значит имеющий ядро). ДНК эукариот линейная (у прокариот ДНК кольцевая и находится в особой области клетки - нуклеоиде, который не отделён мембраной от остальной цитоплазмы). Она связана с белками-гистонами и другими белками хромосом, которых нет у бактерий.В жизненном цикле эукариот обычно присутствуют две ядерные фазы (гаплофаза и диплофаза). Первая фаза характеризуется гаплоидным (одинарным) набором хромосом, далее, сливаясь, две гаплоидные клетки (или два ядра) образуют диплоидную клетку (ядро), содержащую двойной (диплоидный) набор хромосом. Иногда при следующем делении, а чаще спустя несколько делений клетка вновь становится гаплоидной. Такой жизненный цикл и в целом диплоидность для прокариот не характерны.
Третье, пожалуй, самое интересное отличие, - это наличие у эукариотических клеток особых органелл, имеющих свой генетический аппарат, размножающихся делением и окружённых мембраной. Эти органеллы - митохондрии и пластиды. По своему строению и жизнедеятельности они поразительно похожи на бактерий. Это обстоятельство натолкнуло современных учёных на мысль, что подобные организмы являются потомками бактерий, вступившими в симбиотические отношения с эукариотами. Прокариоты характеризуются малым количеством органелл, и ни одна из них не окружена двойной мембраной. В клетках прокариот нет эндоплазматического ретикулума, аппарата Гольджи, лизосом.
Ещё одно важное различие между прокариотами и эукариотами - наличие у эукариот эндоцитоза, в том числе у многих групп - фагоцитоза. Фагоцитозом (дословно «поедание клеткой») называют способность эукариотических клеток захватывать, заключая в мембранный пузырёк, и переваривать самые разные твёрдые частицы. Этот процесс обеспечивает в организме важную защитную функцию. Впервые он был открыт И. И. Мечниковым у морских звёзд. Появление фагоцитоза у эукариот скорее всего связано со средними размерами (далее о размерных различиях написано подробнее). Размеры прокариотических клеток несоизмеримо меньше, и поэтому в процессе эволюционного развития эукариот у них возникла проблема снабжения организма большим количеством пищи. Как следствие среди эукариот появляются первые настоящие, подвижные хищники.
Большинство бактерий имеет клеточную стенку, отличную от эукариотической (далеко не все эукариоты имеют её). У прокариот это прочная структура, состоящая главным образом из муреина (у архей из псевдомуреина). Строение муреина таково, что каждая клетка окружена особым сетчатым мешком, являющимся одной огромной молекулой. Среди эукариот клеточную стенку имеют многие протисты, грибы и растения. У грибов она состоит из хитина и глюканов, у низших растений - из целлюлозы и гликопротеинов, диатомовые водоросли синтезируют клеточную стенку из кремниевых кислот, у высших растений она состоит из целлюлозы, гемицеллюлозы и пектина. Видимо, для более крупных эукариотических клеток стало невозможно создавать клеточную стенку из одной молекулы высокую по прочности. Это обстоятельство могло заставить эукариот использовать иной материал для клеточной стенки. Другое объяснение состоит в том, что общий предок эукариот в связи с переходом к хищничеству утратил клеточную стенку, а затем были утрачены и гены, отвечающие за синтез муреина. При возврате части эукариот к осмотрофному питанию клеточная стенка появилась вновь, но уже на другой биохимической основе.
Разнообразен и обмен веществ у бактерий. Вообще всего выделяют четыре типа питания, и среди бактерий встречаются все. Это фотоавтотрофные, фотогетеротрофные, хемоавтотрофные, хемогетеротрофные (фототрофные используют энергию солнечного света, хемотрофные используют химическую энергию). Эукариоты же либо сами синтезируют энергию из солнечного света, либо используют готовую энергию такого происхождения. Это может быть связано с появлением среди эукариотов хищников, необходимость синтезировать энергию для которых отпала.
Ещё одно отличие - строение жгутиков. У бактерий они тонкие - всего 15–20 нм в диаметре. Это полые нити из белка флагеллина. Строение жгутиков эукариот гораздо сложнее. Они представляют собой вырост клетки, окруженный мембраной, и содержат цитоскелет (аксонему) из девяти пар периферических микротрубочек и двух микротрубочек в центре. В отличие от вращающихся прокариотическох жгутиков жгутики эукариот изгибаются или извиваются.
Две группы рассматриваемых нами организмов, как уже было сказано, сильно отличаются и по своим средним размерам. Диаметр прокариотической клетки составляет обычно 0,5–10 мкм, когда тот же показатель у эукариот составляет 10–100 мкм. Объём такой клетки в 1000–10000 раз больше, чем прокариотической.
Рибосомы прокариот мелкие (70S-типа). Клетки эукариот содержат как более крупные рибосомы 80S-типа, находящиеся в цитоплазме, так и 70s-рибосомы прокариотного типа, расположенные в митохондриях и пластидах.
Видимо, различается и время возникновения этих групп. Первые прокариоты возникли в процессе эволюции около 3,5 млрд. лет назад, от них около 1,2 млрд. лет назад произошли эукариотические организмы.
В настоящее время различают прокариотические (доядерные) и эукариотические (ядерные) клетки. В отличие от прокариотической, эукариотическая клетка имеет ядро, ограниченное оболочкой из двух мембран и большое количество мембранных органелл. К прокариотам принадлежат сине-зеленые водоросли, актиномицеты, бактерии, спирохеты, микоплазмы, риккетсии и хламидии, к эукариотам - большинство водорослей, грибы, лишайники, растения и животные.
Основное отличие прокариотических
клеток от эукариотических заключается
в том, что их ДНК не организована в
хромосомы и не окружена ядерной оболочкой.
Эукариотические клетки устроены значительно
сложнее. Их ДНК, связанная с белком, организована в хромосомы, которые располагаются
в особом образовании, по сути самом крупном
органоиде клетки - ядре. Кроме того, внеядерное
активное содержимое такой клетки ра
и т.д.................
Важнейшая, основополагающая особенность эукариотических клеток связана с расположением генетического аппарата в клетке. Генетический аппарат всех эукариот находится в ядре и защищён ядерной оболочкой. ДНК эукариот линейная (у прокариот ДНК кольцевая и находится в особой области клетки -нуклеоиде, который не отделён мембраной от остальной цитоплазмы). Она связана с белками-гистонами и другими белками хромосом, которых нет у бактерий.
В жизненном цикле эукариот обычно присутствуют две ядерные фазы (гаплофаза и диплофаза). Первая фаза характеризуется гаплоидным (одинарным) набором хромосом, далее, сливаясь, две гаплоидные клетки (или два ядра) образуют диплоидную клетку (ядро), содержащую двойной (диплоидный) набор хромосом. Иногда при следующем делении, а чаще спустя несколько делений клетка вновь становится гаплоидной. Такой жизненный цикл и в целом диплоидность для прокариот не характерны.
Третье отличие - это наличие у эукариотических клеток особых органелл, имеющих свой генетический аппарат, размножающихся делением и окружённых мембраной. Эти органеллы - митохондрии и пластиды. По своему строению и жизнедеятельности они поразительно похожи на бактерий.. Прокариоты характеризуются малым количеством органелл, и ни одна из них не окружена двойной мембраной.
Ещё одно важное различие между прокариотами и эукариотами - наличие у эукариот эндоцитоза, в том числе у многих групп - фагоцитоза. Фагоцитозом называют способность эукариотических клеток захватывать, заключая в мембранный пузырёк, и переваривать самые разные твёрдые частиц. Размеры прокариотических клеток несоизмеримо меньше, и поэтому в процессе эволюционного развития эукариот у них возникла проблема снабжения организма большим количеством пищи. Как следствие среди эукариот появляются первые настоящие, подвижные хищники.
Большинство бактерий имеет клеточную стенку, отличную от эукариотической. Строение муреина таково, что каждая клетка окружена особым сетчатым мешком, являющимся одной огромной молекулой. Среди эукариот клеточную стенку имеют многие протисты, грибы и растения. У грибов она состоит из хитина и глюканов, у низших растений - из целлюлозы и гликопротеинов, диатомовые водоросли синтезируют клеточную стенку из кремниевых кислот, у высших растений она состоит из целлюлозы, гемицеллюлозы и пектина. Видимо, для более крупных эукариотических клеток стало невозможно создавать клеточную стенку из одной молекулы высокую по прочности. Это обстоятельство могло заставить эукариот использовать иной материал для клеточной стенки. Другое объяснение состоит в том, что общий предок эукариот в связи с переходом к хищничеству утратил клеточную стенку, а затем были утрачены и гены, отвечающие за синтез муреина. При возврате части эукариот к осмотрофному питанию клеточная стенка появилась вновь, но уже на другой биохимической основе.
Разнообразен и обмен веществ у бактерий. Вообще всего выделяют четыре типа питания, и среди бактерий встречаются все. Это фотоавтотрофные, фотогетеротрофные, хемоавтотрофные, хемогетеротрофные. Эукариоты же либо сами синтезируют энергию из солнечного света, либо используют готовую энергию такого происхождения. Это может быть связано с появлением среди эукариотов хищников, необходимость синтезировать энергию для которых отпала.
Ещё одно отличие - строение жгутиков. У бактерий жгутиками являются полые нити диаметром 15–20 нм из белка флагеллина. Строение жгутиков эукариот гораздо сложнее. Они представляют собой вырост клетки, окруженный мембраной, и содержат цитоскелет (аксонему) из девяти пар периферических микротрубочек и двух микротрубочек в центре. В отличие от вращающихся прокариотическох жгутиков жгутики эукариот изгибаются или извиваются.
Две группы рассматриваемых нами организмов, как уже было сказано, сильно отличаются и по своим средним размерам. Диаметр прокариотической клетки составляет обычно 0,5–10 мкм, когда тот же показатель у эукариот составляет 10–100 мкм. Объём такой клетки в 1000–10000 раз больше, чем прокариотической.
Клетки эукариот содержат как более крупные рибосомы. Первые прокариоты возникли в процессе эволюции около 3,5 млрд. лет назад, от них около 1,2 млрд. лет назад произошли эукариотические организмы.
8.Бактерии - это одноклеточные организмы растительного происхождения, но лишенные хлорофилла. Они относятся к прокариотам, видны в световой микроскоп, размеры их измеряются в микрометрах. Бактерии растут на искусственных питательных средах, размножение происходит бинарным делением.
Бактерии делятся
вызывают болезни челове- вызывают заболевания не вызывают забо-
ка, животных, растений при определенных усло- леваний
Морфология - это форма, размер бактерий, расположение клеток в препарате. Различают три морфологических формы бактерий:
Различают три морфологических формы бактерий:
1) кокки 2) палочки 3) извитые
1. Кокки: форма - круглая
размер - мелкие
расположение в препаратах - 6 разновидностей:
а) микрококки б) диплококки в) тетракокки
гонококки пневмококки
г) сарцины в) стафилококки д) стрептококки
2. Палочковидные :
· форма - цилиндр
· размер: длина : толщина :
Крупные - толстые
Средних размеров - тонкие
· концы палочек – закругленные (кишечная палочка)
Прямые (сибиреязвенная палочка)
В виде утолщения (дифтерийная палочка)
· расположение - беспорядочное
В цепочку (стрептобактерии)
Попарно (диплобактерии)
В виде римских цифр II, V, X и т.д.
3. Извитые : форма – спиралевидная:
1. спириллы,
2. кампилобактерии
Бактериальная клетка состоит из клеточной стенки, цитоплазматической мембраны, цитоплазмы с включениями и ядерного аппарата, называемого нуклеоидом. Имеются другие структуры: мезосома, хроматофоры, тилакоиды, вакуоли, включения полисахаридов, жировые капельки, капсула (микрокапсула, слизь), жгутики, пили. Некоторые бактерии способны образовывать споры.
Структуру и морфологию бактерий изучают с помощью различных методов микроскопии: световой, фазово-контрастной, интерференционной, темнопольной, люминесцентной и электронной.
Клеточная стенка
В клеточной стенки грамположительных бактерий содержится небольшое количество полисахаридов, липидов, белков. Основным компонентом клеточной стенки этих бактерий является многослойный пептидогликан (муреин, мукопептид), составляющий 40-90% массы клеточной стенки. С пептидогликаном клеточной стенки грамположительных бактерий ковалентно связаны тейхоевые кислоты (от греч. teichos - стенка).
В состав клеточной стенки грамотрицательных бактерий входит наружная мембрана, связанная посредством липопротеина с подлежащим слоем пептидогликана. На ультратонких срезах бактерий наружная мембрана имеет вид волнообразной трехслойной структуры, сходной с внутренней мембраной, которую называют цитоплазматической. Основным компонентом этих мембран является бимолекулярный (двойной) слой липидов. Внутренний слой наружной мембраны представлен фосфолипидами, а в наружном слое расположен липополисахарид (ЛПС). Липополисахарид наружной мембраны состоит из трех фрагментов: липида А - консервативной структуры, практически одинаковой у грамотрицательных бактерий; ядра, или стержневой, коровой части (лат. core - ядро), относительно консервативной олигосахаридной структуры (наиболее постоянной частью ядра ЛПС является кетодезоксиоктоновая кислота); высоковариабельнои О-специфической цепи полисахарида, образованной повторяющимися идентичными олигосахаридными последовательностями (О-антиген). Белки матрикса наружной мембраны пронизывают ее таким образом, что молекулы белка, называемые поринами, окаймляют гидрофильные поры, через которые проходят вода и мелкие гидрофильные молекулы.
При нарушении синтеза клеточной стенки бактерий под влиянием лизоцима,
пенициллина, защитных факторов организма образуются клетки с измененной (часто шаровидной) формой: протопласты - бактерии, полностью лишенные клеточной стенки; сферопласты - бактерии с частично сохранившейся клеточной стенкой. Бактерии сферо- или протопластного типа, утратившие способность к синтезу пептидогликана под влиянием антибиотиков или других факторов и способные размножаться, называются L-формами.
Они представляют собой осмотически чувствительные, шаровидные, колбовидные клетки различной величины, в том числе и проходящие через бактериальные фильтры. Некоторые L-формы (нестабильные) при удалении фактора, приведшего к изменениям бактерий, могут реверсировать, «возвращаясь» в исходную бактериальную клетку.
Между наружной и цитоплазматической мембранами находится периплазматическое пространство, или периплазма, содержащая ферменты (протеазы, липазы, фосфатазы, нуклеазы, бета-лактомазы) и компоненты транспортных систем.
Статьи по теме