Клеточная стенка растений состоит из целлюлозы. Клеточная стенка и ее роль в жизнедеятельности растительной клетки
Наименьшей частью организма является клетка, она способна существовать самостоятельно и имеет все признаки живого организма. В данной статье мы узнаем, какое строение имеет растительная клетка, кратко расскажем об её функциях и особенностях.
Строение клетки растения
В природе существуют как одноклеточные растения, так и многоклеточные. Например, в подводном мире можно встретить одноклеточные водоросли, которые имеют все функции присущие живому организму.
Многоклеточная особь - это не просто набор клеток, а единый организм, способный образовывать различные ткани, органы, которые взаимодействуют друг с другом.
Строение растительной клетки у всех растений одинаковое и состоит из одних и тех же компонентов. Её состав следующий:
- оболочка (пластинка, межклетник, плазмодесмы и плазмолеммы, тонопласт);
- вакуоли;
- цитоплазма (митохондрии; хлоропласты и другие органоиды);
- ядро (ядерная оболочка, ядрышко, хроматин).

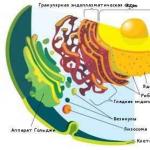
Рис. 1. Строение клетки растения.
В отличие от животной, растительная клетка имеет особую целлюлозную оболочку, вакуоль и пластиды.
Изучение строения и функций растительной клетки показало, что:
ТОП-4 статьи которые читают вместе с этой
- самой значительной частью в организме является ядро , которое отвечает за все происходящие процессы. Оно содержит наследственную информацию, которая передаётся из поколения в поколение. От других органоидов отделяет ядро ядерная оболочка;
- бесцветное вязкое вещество, которое наполняет клетку, называется цитоплазмой . Именно в ней находятся все органоиды;
- под клеточной стенкой находится мембрана (тонопласт) , которая отвечает за обмен веществ. Это тоненькая плёнка, отделяющая оболочку от цитоплазмы;
- клеточная стенка достаточно прочная, так как в её состав входит целлюлоза. Поэтому функциями стенки является защита и придача формы;
- маленькими составными компонентами являются пластиды
.
Они могут быть цветными или бесцветными. Так, например, хлоропласты имеют зелёный цвет, именно в них происходит процесс фотосинтеза;
- внутренняя полость, заполненная соком, называется вакуолью . Размер её зависит от возраста организма: чем он старше, тем больше вакуоль. В состав сока входит водный раствор минеральных солей и органических веществ. Он содержит различные сахара, ферменты, минеральные кислоты и соли, белки и пигменты;
- митохондрии способны передвигаться вместе с цитоплазмой, их основная роль - обмен веществ. Именно здесь происходит процесс дыхания и образования АТФ;
- аппарат Гольджи может иметь различные формы (диски, палочки, зёрнышки). Его роль - накопление и выведение ненужных веществ;
- рибосомы синтезируют белок. Находятся они в цитоплазме, ядре, митохондриях, пластидах.
- в отличие от других живых организмов, растения имеют вакуоль, которая хранит все питательные и полезные вещества, расщепляет отжившие старые органеллы и белки;
- клеточная стенка по своему составу отличается от грибного хитина и стенок бактерий. В её состав входит целлюлоза, пектин и лигнин;
- связь между клетками осуществляется при помощи плазмодесм – так называемые поры в клеточной стенке;
- пластиды имеются только в растительном организме. Помимо хлоропластов это могут быть лейкопласты, которые делятся на два вида: одни из них запасают жиры, другие - крахмал. А также хромопласты, которые синтезируют и хранят пигменты;
- в отличие от животного организма, у растительной клетки нет центриолей.

Рис. 2. Изменения размера вакуоли при росте растения.
Клеточное строение растений учёные открыли ещё в XVII веке. Клетки апельсиновой мякоти видны невооружённым глазом, но чаще всего рассмотреть растительный организм можно под микроскопом.

Рис. 3. Строение аппарата Гольджи.
Особенности растительного организма
Исследование разнообразия царства растений выявило такие особенности:
Что мы узнали?
Будучи самой маленькой частью всего организма, клетка может существовать самостоятельно. Она обеспечивает работу различных тканей и жизненно важных органов. Отличительными компонентами от других особей живой природы является строение клеточной стенки, наличие пластид и вакуолей. Каждый органоид имеет свои функции, без выполнения которых невозможно функционирование всего организма в целом.
Тест по теме
Оценка доклада
Средняя оценка: 4.3 . Всего получено оценок: 1130.
Расположенная снаружи от цитоплазматической мембраны и выполняющая структурные, защитные и транспортные функции. Обнаруживается у большинства бактерий , архей , грибов и растений . Животные и многие простейшие не имеют клеточной стенки.
Клеточные стенки прокариот
Клеточные стенки бактерий состоят из пептидогликана (муреина) и бывают двух типов: грамположительного и грамотрицательного . Клеточная стенка грамположительного типа состоит исключительно из толстого слоя пептидогликана, плотно прилегающего к клеточной мембране и пронизанного тейхоевыми и липотейхоевыми кислотами . При грамотрицательном типе слой пептидогликана существенно тоньше, между ним и плазматической мембраной находится периплазматическое пространство , а снаружи клетка окружена ещё одной мембраной , представленной т. н. липополисахаридом и являющаяся пирогенным эндотоксином грамотрицательных бактерий.
Клеточные стенки грибов
Клеточные стенки водорослей
Клеточные стенки высших растений
Важнейшей отличительной особенностью растительной клетки является наличие прочной клеточной стенки, основным компонентом которой является целлюлоза. Клеточная стенка высших растений представляет собой сложноорганизованный, преимущественно полимерный, внеклеточный матрикс, окружающий каждую клетку. Растительная клетка, лишённая клеточной стенки, обозначается термином протопласт . В клеточных стенках растений существуют углубления - поры, через которые проходят цитоплазматические канальца - плазмодесмы , осуществляющие контакт соседних клеток и обмен веществами между ними.
Химический состав и пространственная организация полимеров клеточной стенки отличаются у разных видов, клеток разных тканей одного растения и иногда у разных частей стенки вокруг одного протопласта.
Кроме того, строение клеточной стенки изменяется в онтогенезе растительного организма. Первичная клеточная стенка формируется при делении и сохраняется во время роста клетки. Формирование вторичной клеточной стенки происходит с внутренней стороны от первичной стенки и связано с окончанием роста и специализацией (дифференцировкой) клеток растения. Снаружи от первичной клеточной стенки, между первичными стенками двух соседних клеток, располагается срединная пластинка (состоит преимущественно из кальциевых и магниевых солей пектиновых веществ).
Первичная клеточная стенка высших растений состоит из трёх взаимодействующих, но структурно независимых трехмерных сетей полимеров. Основная сеть состоит из фибрилл целлюлозы и связывающих их гемицеллюлоз (или сшивочных гликанов). Вторая сеть состоит из пектиновых веществ . Третья сеть представлена, как правило, структурными белками клеточной стенки. Также следует отметить, что у растений клады commelinids (группа в системах APG) и у представителей семейства Маревые первичная клеточная стенка содержит значительное количество ароматических веществ (гидроксикоричные кислоты , главным образом феруловая и п -кумаровая). При этом у представителей клады commelinids гидроксикоричные кислоты присоединяются к сшивочным гликанам (к глюкуроноарабиноксиланам), а у семейства Маревые к пектиновым веществам (к рамногалактуронанам I).
Растительные клеточные стенки выполняют целый ряд функций: они обеспечивают жёсткость клетки для структурной и механической поддержки, придают форму клетке, направление её роста и в конечном счете морфологию всему растению. Клеточная стенка также противодействует тургору, то есть осмотическому давлению, когда дополнительное количество воды поступает в растения. Клеточные стенки защищают от патогенов, проникающих из окружающей среды, и запасают углеводы для растения.
Материал для построения клеточной стенки секретирует сам заключенный в ней – протопласт. Клеточная стенка, отлагающаяся во время деления клеток растения, называется первичной клеточной стенкой. Позже в результате утолщения она может превратиться во вторичную клеточную стенку.
Первичная клеточная стенка состоит из целлюлозных мицелл, которые образуют микрофибриллы, погруженных в матрикс, в состав которого входят сложные полисахариды.
Целлюлоза представляет собой полисахарид (полимер глюкозы). В ней заключено около 50 % углерода, находящегося в растениях. 20–40 % материала клеточной стенки составляет целлюлоза. Целлюлозные волокна представляют собой длинные цепи – приблизительно из 10 000 остатков глюкозы. Из каждой цепи выступает наружу множество ОН-групп. Эти группы направлены во все стороны и образуют водородные связи с соседними цепями, что обеспечивает жесткое поперечное сшивание всех цепей. Цепи объединены друг с другом, образуя микрофибриллы, в которых происходит образование пространственных решеток, кристаллоподобных участков, так называемых мицелл (рис. 1).
Рис. 1.
1 – микрофибрилла; 2 – мицелла; 3 – межмицеллярное пространство (диаметр 1 нм); 4 – межмикрофибиллярное пространство (диаметр ~ 10 нм)
Особо важное значение для той роли, которую выполняет целлюлоза в клеточных стенках имеет ее волокнистое строение и высокая прочность на разрыв, сравнимая с прочностью стали.
Микрофибриллы, погруженные в матрикс, образуют каркас клеточной стенки. Матрикс клеточной стенки состоит из полисахаридов, которые для удобства делят на пектины и гемицеллюлозы в зависимости от их растворимости в различных растворителях.
Гемицеллюлозы – это смешанная группа полисахаридов, растворимая в щелочах (к ним относятся полимеры ксилозы, галактозы, маннозы, глюкозы и глюкоманнозы). У гемицеллюлозы, как и у целлюлозы, молекулы имеют форму цепи, однако их цепи короче, менее упорядочены и сильнее разветвлены.
Молекулы пектина представляют собой цепочки полигалактуроновой кислоты, свободные карбоксильные группы которой составляют резервуар фиксированных отрицательных зарядов в клеточных стенках.
Отметим также, что размер межмицеллярного и межфибриллярного пространства (пор) составляет соответственно 1 и 10 нм, что значительно превышает размер многих растворенных веществ, транспортируемых в растениях.
Среди вещества матрикса клеточных стенок имеются и белки . Это гликопротеиды, белковая фракция которых сильно обогащена аминокислотой оксипролином. Структурный белок клеточной стенки получил название экстенсина.
Таким образом, свободные карбоксильные группы полисахаридов и белков обеспечивают большое число отрицательных фиксированных зарядов в клеточных стенках. Свободные аминогруппы белков ответственны за меньшее количество положительных зарядов. Заряженные группы матрикса образуют доннановскую систему, играющую важную роль в процессе транспорта ионов в тканях растений. Клеточные стенки гидратированы: 60–70 % их массы обычно составляет вода. По свободному пространству клеточной стенки вода перемещается беспрепятственно. Присутствие воды оказывает влияние на химические и физические свойства полисахаридов клеточной стенки.
У некоторых клеток, например у клеток мезофилла листа, на всем протяжении их жизни имеется только первичная клеточная стенка. Однако у большинства клеток на внутреннюю поверхность первичной стенки отлагаются дополнительные слои целлюлозы, т. е. образуется вторичная клеточная стенка. Обычно это происходит после того, как клетка достигнет своего максимального размера, и лишь немногие клетки, например клетки колленхимы, продолжают рост во время этой фазы. Вторичное утолщение клеточных стенок растения не следует путать со вторичным утолщением (вторичным ростом) самого растения, т. е. увеличением толщины ствола в результате добавления новых клеток.
Самый внутренний, очень тонкий последний слой вторичной стенки вследствие особенностей структуры и состава принято называть третичной стенкой. Наряду с целлюлозой и пектином в этом слое имеется много гемоцеллюлоз: в третичной стенке микрофибриллы, по-видимому, образуют сетчатую структуру. Обращенная внутрь клетки поверхность третичной стенки обычно покрыта субмикроскопическими бородавчатыми выростами.
Некоторые клетки, такие, как трахеальные элементы ксилемы и клетки склеренхимы, претерпевают интенсивную лигнификацию (одревеснение); при этом все слои целлюлозы (первичный и вторичные) пропитываются лигнином – сложным полимерным веществом, не относящимся к полисахаридам. В клетках протоксилемы отложения лигнина имеют кольцевую, спиральную или сетчатую форму. В других случаях лигнификация бывает сплошной, если не считать так называемых поровых полей, т. е. тех участков в первичной клеточной стенке, через которые осуществляется контакт между соседними клетками при помощи плазмодесм. Лигнин скрепляет целлюлозные волокна и удерживает их на месте. Он действует как очень твердый и жесткий матрикс, усиливающий прочность клеточных стенок на растяжение и в особенности на сжатие. Он же обеспечивает клеткам дополнительную защиту от неблагоприятных физических и химических воздействий. В ряде случаев клетки пропитываются суберином или покрыты восковым налетом – кутикулой.
Клеточная стенка, как упоминалось, пронизана порами, сквозь которые проходят плазмодесмы. Плазмодесма – это не просто нить цитоплазмы, соединяющая протопласты соседних клеток, а органелла имеющая сложное строение. Снаружи плазмодесма покрыта плазмалеммой, которая переходит из одной клетки в другую. Внутри плазмодесмы находится канал (десмотубола), соединяющий эндоплазматическую сеть соседних клеток. Пора, находящаяся в клеточной стенке, в средней части расширена, а на концах сужена, поэтому на концах плазмалемма почти вплотную подходит к десмотуболе. На 1 мм 2 клеточной стенки приходится от 100 тыс. до 50 млн. плазмодесм.
Совокупность протопластов всех клеток, соединенных плазмодесмами, получила название симпласта.
Сравнительно недавно были открыты эктодесмы – очень тонкие цитоплазматические выросты, пронизывающие наружные стенки эпидермальных клеток листа и доходящие до кутикулы. Они подвижны: могут укорачиваться и удлиняться. Эктодесмы, вероятно, выполняют в многоклеточном организме роль рецепторов: воспринимают разного рода сигналы об изменениях в окружающей среде и передают их затем симпласту.
Основные функции клеточной стенки (оболочки) следующие:
1. Оболочки обеспечивают отдельным клеткам и растению в целом механическую прочность и опору. В некоторых тканях прочность усиливается благодаря интенсивной лигнификации клеточных стенок (небольшое количество лигнина присутствует во всех клеточных стенках).
2. Относительная жесткость клеточных стенок и сопротивление растяжению обуславливают и тургесцентностьклеток, когда в них осмотическим путем поступает вода. Это усиливает опорную функцию во всех растениях и служит единственным источником опоры для травянистых растений и для таких органов, как листья, т. е. там, где отсутствует вторичный рост. Клеточные стенки также предохраняют клетки от разрыва в гипотонической среде.
3. Ориентация целлюлозных микрофибрилл ограничивает, и в известной мере, регулирует как рост, так и форму клеток, поскольку от расположения этих микрофибрилл зависит способность клеток к растяжению. Если, например, микрофибриллы располагаются поперек клетки, окружая ее как бы обручами, то клетка, в которую путем осмоса поступает вода, будет растягиваться в продольном направлении.
4. Система связанных друг с другом клеточных стенок (апопласт) служит путем, по которому передвигается вода и минеральные вещества. Клеточные стенки скреплены между собой с помощью срединных пластинок. В стенках имеются небольшие поры, сквозь которые проходят цитоплазматические тяжи – плазмодесмы. Плазмодесмы объединяют все протопласты в единую систему – симпласт.
5. Наружные клеточные стенки эпидермальных клеток покрываются особой пленкой – кутикулой, состоящей из воскообразного вещества кутина, что снижает потерю воды и уменьшает риск проникновения в растение болезнетворных организмов. В пробковой ткани клеточные стенки по завершению вторичного роста пропитываются суберином, выполняющим сходную функцию.
6. Клеточные стенки сосудов ксилемы, трахеид и ситовидных трубок приспособлены для дальнего транспорта веществ по растению.
7. Стенки клеток эндодермы корня пропитаны суберином и поэтому служат барьером на пути движения воды и минеральных веществ.
8. У некоторых клеток их видоизмененные стенки хранят запасы питательных веществ. Таким способом, например, запасаются гемицеллюлозы в некоторых семенах.
9. У передаточных клеток (клетки-спутники) площадь поверхности клеточных стенок увеличена и соответственно увеличена площадь поверхности плазмалеммы, что повышает эффективность переноса веществ.
10. Благодаря своим катионобменным свойствам клеточная стенка служит резервуаром, накапливающим катионы.
Клеточная стенка бактерий - это тонкая бесцветная структура, покрывающая клетку снаружи. У большинства бактерий она невидима в обыкновенный микроскоп без специальной обработки. Однако у крупных форм, например у серобактерии Beg. mirabilis, стенка заметна отчетливо. При явлении плазмолиза, который наступает при помещении клеток в 1-2 %-ный гипертонический раствор NaCl или раствор глюкозы, контуры стенки приобретают четкость и она хорошо видна при фазово-контрастной микроскопии.
Стенка бактериальной клетки составляет до 50 % сухой массы организма, толщина ее колеблется в пределах 20-80 нм. Клеточная стенка - плотная ригидная структура. Она обладает эластичностью и достаточной механической прочностью, выдерживает внутриклеточное осмотическое давление, достигающее 10-30 атм.
Химический состав клеточных стенок различных видов бактерий неодинаков, довольно сложен и отличает их не только от клеток растений и животных, но и друг от друга.
Основным компонентом клеточной оболочки высших растений и водорослей является целлюлоза. Из целлюлозы состоят, например, микрофибриллы большинства водорослей - до 50-80 % сухой массы оболочки клетки. В микрофибриллах клеточных оболочек большинства мицелиальных грибов преобладает хитин - полимер N-ацетилглюкозамина.
Совершенно иной химический состав имеют клеточные стенки бактерий. Такие соединения как целлюлоза и хитин не характерны для них. Правда, некоторые виды бактерий способны синтезировать целлюлозу и компоненты хитина. Так, у Sarcina ventriculi целлюлоза составляет толстый внешний слой клеточной стенки. Помимо Acetobacter xylinum, это единственный представитель прокариот, синтезирующий данный полимер. Компонент хитина ацетилглюкозамин обнаруживается у всех видов бактерий, за исключением некоторых архебакгерий.
В клеточных стенках бактерий содержится два класса новых, необычных соединений, присущих только прокариотам. Это пептидогликан и тейхоевые кислоты.
Пептидогликаны и тейхоевые кислоты. Пептидогликан, или муреин (от лат. myrus - стенка) представляет собой гетерополимер, состоящий из цепочек чередующихся остатков N-ацетилглюкозамина и N-ацетилмурамовой кислоты (эфир молочной кислоты и N-ацетилглюкозамина), соединенных р-1,4-гликозидной связью. К карбоксильной группе мурамовой кислоты присоединен пептид, включающий чаще всего четыре аминокислоты - тетрапептид. Аминокислотный состав пептида различных видов бактерий не одинаков: у Staph, aureus содержится а-лизин, у Е. сой - мезо-диаминопимелиновая кислота, у Corynebacterium - 2-4-диаминомасляная (рис. 3.11).
Рис. 3.11. Структура пептидогликана стафилококка:
1- N-ацетилмурамовая кислота; 2 - N-ацетилглюко- замин; 3 - тетрапептид; 4 - глидоновый мостик
На основании аминокислотного состава пептидов и соединяющих их мостиков различают ряд подгрупп пептидогликана. Особенностью пептидной части этого полимера является наличие D-аминокислот (в белках они не встречаются) и высокое содержание диаминокислот. Обе аминогруппы, входящие в состав муреина диаминокислоты, участвуют в образовании пептидных связей - с D-аланином и аминокислотным мостиком. Посредством мостиков осуществляются поперечные сшивки пептидогликановых цепей. В итоге формируется гигантская молекула, напоминающая по виду мешок, состоящая из сети полисахаридных цепей, связанных множеством поперечных пептидных связей. За счет образования поперечных сшивок обеспечивается жесткая трехмерная пространственная организация молекулы, обусловливающая механическую прочность и ригидность клеточной стенки.
Пептидогликан чувствителен к литическому действию лизоцима, который расщепляет p-l-4-гликозидные связи между N-ацетилглюкозамином и N-ацетилмурамовой кислотой. Обработка бактерий лизоцимом приводит к разрушению сформированной клеточной стенки. Ингибитором синтеза пептидогликана является ряд антибиотиков: пенициллин, цефалоспорин, бацитрацин,
ванкомицин. К примеру, пенициллин подавляет активность фермента транспептидазы, катализирующего образование поперечных сшивок между образующимися цепями пептидогликана. Не сшитый полимер не используется для образования клеточной стенки бактерий.
Тейхоевые кислоты (от греч. «тейхос» - стенка) представляют собой растворимые в воде полимеры, состоящие из остатков трехатомного спирта глицерола или пятиатомного - рибитола, которые соединены друг с другом фосфодиэфирными связями (рис. 3.12). Цепи тейхоевых кислот могут содержать от 10 до 50 остатков спирта. Большинство тейхоевых кислот включают значительное количество D-аланина, аминогруппы которого придают тейхоевым кислотам амфотерные свойства. Кроме D-аланина свободные гидроксильные группы спиртов могут быть замещены глюкозой, N-ацетилглюкозамином, галактозой. Наличие свободных гидро-
ксилов фосфорной кислоты обусловливает сродство тейхоевых кис лот к двухвалентным катионам.

Рис.3.12. Структура тейхоевых кислот клеточной стенки:
а -глицеролтейхоевая; б - рибитолтеихоевая
Клетки одного штамма бактерий, как правило, содержат тейхоевую кислоту только одного типа: рибитолтейхоевую или глицеролтейхоевую. Эти уникальные соединения содержатся в клеточных стенках только грамположительных бактерий, где они прочно связаны с пептидогликаном. Так как тейхоевые кислоты представляют собой длинные линейные молекулы, они могут проходить через весь пептидогликановый слой до наружной части клетки и играть роль поверхностных антигенов, обусловливая, таким образом, антигенную специфичность клеточной поверхности бактерий. Кроме того, создавая в клеточной стенке высокую плотность строго ориентированных зарядов, тейхоевые кислоты оказывают влияние на проникновение ионов в клетку, обеспечивая высокую плотность двухвалентных катионов в области цитоплазматической мембраны. Это благоприятствует поддержанию физической целостности мембраны и ее связи с рибосомами.
У некоторых бактерий тейхоевые кислоты участвуют в регуляции активности автолитических ферментов, осуществляющих
в определенных условиях гидролиз муреина собственной клетки. Так, у пневмококков тейхоевые кислоты ингибируют действие литических ферментов клетки путем связывания с ними. Нарушение этой связи приводит клетки к лизису.
Пептидогликан является основным структурным компонентом клеточных стенок почти всех прокариот, за исключением архебактерий, у которых он либо совсем отсутствует, либо имеет иной химический состав. Например, у метанобразующих бактерий пептидогликан содержит вместо муреиновой кислоты талозоминуроновую, а пептидная часть не содержит D-аминокислот, состоит только из а-форм.
В зависимости от химического состава и структуры клеточной стенки все бактерии разделяют на грамположительные и грамотрицательные. Это основано на способности их окрашиваться фиолетовыми красителями трифенилметанового ряда кристаллвиолетом или генцианвиолетом - и не обесцвечиваться нейтральными растворителями - спиртом, ацетоном. Этот метод окраски введен впервые в 1884 г. датским врачом Христианом Грамом и окраска по Граму используется как важнейший таксономический признак бактерий. Сущность его состоит в следующем. Фиксированные клетки окрашиваются кристаллвиолетом или генцианвиолетом, затем протравливаются 30 с раствором Люголя (1 + KI), промываются спиртом, водой и докрашиваются 1 %-ным водным фуксином. Грамположительные бактерии приобретают синий цвет, грамотрицательные - красный.
По структуре и химическому составу клеточной стенки грамположительные бактерии существенно отличаются от грамотрицательных (табл. 2).
У грамположительных бактерий клеточная стенка представляет собой гомогенный электронно-плотный слой толщиной 20 - 80 нм. Основную массу (50-90 % сухого вещества) составляет пептидогликан, образующий ригидный толстый слой. Он плотно прилегает к ЦПМ. Пептидогликановый слой пронизан тейхоевыми кислотами, которые могут выходить на поверхность клеточной стенки. Кроме этих основных полимеров в клеточных стенках грамположительных бактерий содержатся в небольших количествах липиды, полисахариды, белки. Липиды и
полисахариды ковалентно связываются с пептидогликаном, образуя сложную, механически прочную структуру.
Таблица 2
Характеристика химического состава клеточных стенок бактерий
точная стенка грамотрицательных бактерий более тонкая (10-15 нм) и многослойная (рис.3.13). Внутренний слой представлен пептидогликаном, содержание которого значительно меньше (1-10 %), чем в стенках грамположительных бактерий. Толщина данного слоя 2-3 нм. Наружный слой более рыхлый и толстый - 8-10 нм, имеет сложный химический состав. В нем обнаружены белки, фосфолипиды и липополисахариды, расположенные мозаично. По структуре и химическому составу этот слой имеет сходство с цитоплазматической мембраной. Он получил название наружной мембраны и имеется только у грамотрицательных бактерий.
Наружная мембрана является дополнительным барьером, препятствующим проникновению в клетку крупных молекул. Так, она препятствует поступлению в клетку антибиотиков, в частности пенициллина, актиномицина Д. Вполне возможно, что по этой причине грамотрицательные бактерии менее чувствительны к антибиотикам, чем грамположительные.
Липополисахариды наружной мембраны определяют антигенную специфичность бактерий, а также служат рецепторами для адсорбции фагов.
Белки наружной мембраны выполняют разные функции. Одни из них, так называемые белки матриксапорины, формируют в
мембране гидрофильные поры, через которые осуществляется диффузия аминокислот, небольших олигосахаридов и пептидов (молекулярная масса от 600 до 900 Да6). Транспорт веществ через поры, образованные поринами, лишен специфичности. Порины являются также рецепторами для фагов и колицинов.

Вторая группа белков - минорные белки, как и предыдущая группа, выполняют транспортные и рецепторные функции. Важная роль отводится им в транспорте железосодержащих соединений в клетке разных видов грамотрицательных бактерий.
Таким образом, структура клеточной стенки грамотрицательных бактерий намного сложнее, чем грамположительных. Структурные особенности и химический состав клеточных стенок лежат в основе механизма окрашиваемости бактерий по Г раму.
Да - дальтон, или единица аггомной массы, равен 1,66033 х 10 1 кг.
Ответственность за окраску по Граму несут муреин и частично липиды, оказывающие влияние на проницаемость клеточной стенки. Обработка бактерий спиртом вызывает разбухание муреина и уменьшение диаметра пор клеточной стенки, что в целом приводит к снижению ее проницаемости. Так как грамположительные бактерии характеризуются высоким содержанием муреина, то в результате обработки спиртом стенки их становятся почти непроницаемыми для красителей и вымывание краски не происходит. У грамотрицательных слой муреина тонкий и не играет существенной роли в проницаемости стенки. Кроме того, проницаемость клеточной стенки у грамположительных бактерий увеличивается за счет растворения и вымывания липидов спиртом, содержание которых довольно высокое (до 22 %), и к тому же они хорошо растворяются в нейтральных органических растворителях. Все это способствует обесцвечиванию клетки. Доказательством того, что в окраске по Граму основную роль играет клеточная стенка, является тот факт, что при удалении ее с окрашенных клеток протопласты грамположительных бактерий при промывании спиртом обесцвечиваются, превращаясь в грамотрицательные. Следовательно, окрашенный комплекс удерживает клеточная стенка.
Клеточная стенка у грамотрицательных бактерий отделена от цитоплазматической мембраны электронно-прозрачным промежутком, получившим название периплазматического пространства, или периплазма. В нем содержатся кроме тонкого слоя муреина (2-3 нм) специфические белки, так называемые связывающие, или транспортные белки. Это водорастворимые белки, обладающие высоким сродством к определенным питательным субстратам - аминокислотам, сахарам, неорганическим ионам. Они являются составной частью систем активного транспорта, но самостоятельно осуществлять этот процесс они не могут и функционируют только в сочетании со специфическими пермеазами, локализованными в цитоплазматической мембране. Транспортные белки связывают соответствующие субстраты и переносят их от внешней мембраны к цитоплазматической. В периплазматическом пространстве содержится также ряд гидролитических ферментов - нуклеазы, щелочная и кислая фосфатазы, пенициллианаза. У грамположительных бактерий эти ферменты являются типичными экзоферментами, у грамотрицательных выход их из клеток задерживается наружной мембраной, которая является барьером для белков и некоторых других соединений. Наличие в периплазме ферментов позволяет клетке использовать более широкий круг веществ, поступающих извне. Так как данные ферменты изолированы от цитоплазмы, то содержание их не угрожает содержимому клетки подвергнуться автолизу, или самоперевариванию.
Важнейшие функции клеточной стенки заключаются в следующем. Она обеспечивает клетке определенное постоянство формы, защищает содержимое от ее внешних воздействий, определяет способность к адсорбции фагов, так как на ее поверхности расположены фагочувствительные рецепторы, играет важную роль и в реакции иммунитета. Установлено, что между фагоцитарной активностью лейкоцитов и поверхностной структурой бактериальных клеток существует определенная зависимость. Особенности структуры клеточной стенки определяют чувствительность бактерий к повреждающему действию сыворотки крови и форменных элементов.
Таким образом, клеточная стенка бактерий - сложная полифункциональная система, обладающая необходимыми реологическими свойствами (упругость, пластичность, прочность) и обеспечивающая анатомическую целостность клетки, геометрическую форму ее и контакт с внешней средой.
Растительные клетки, подобно клеткам прокариот и грибов, заключены в сравнительно жесткую клеточную стенку. Материал для построения этой клеточной стенки секретирует сама заключенная в ней живая клетка (протопласт). По своему химическому составу клеточные стенки растений отличаются от клеточных стенок прокариот и грибов (табл. 2.1), но этим структурам свойственны некоторые общие функции, а именно функции опоры и защиты; кроме того, и те и другие ограничивают подвижность клеток. Клеточная стенка, отлагающаяся во время деления клеток растения, называется первичной клеточной стенкой. Позже в результате утолщения она может превратиться во вторичную клеточную стенку. В этом разделе мы опишем процесс образования первичной клеточной стенки. На рис. 7.21 воспроизведена электронная микрофотография, на которой можно видеть одну из ранних стадий этого процесса.
Строение клеточной стенки
Первичная клеточная стенка состоит из целлюлозных микрофибрилл, погруженных в матрикс, в состав которого входят сложные полисахариды. Целлюлоза тоже представляет собой полисахарид (ее химическое строение описано в разд. 5.2.3). Особо важное значение для той роли, которую целлюлоза выполняет в клеточных стенках, имеют ее волокнистое строение и высокая прочность на разрыв, сравнимая с прочностью стали. Отдельные молекулы целлюлозы - это длинные полисахаридные цепи. Множество таких молекул, сшитых друг с другом поперечными водородными связями, собраны в прочные пучки, называемые микрофибриллами . Погруженные в матрикс микрофибриллы образуют каркас клеточной стенки. Матрикс клеточной стенки состоит из полисахаридов, которые для удобства описания делят обычно на пектины и гемицеллюлозы в зависимости от их растворимости в различных растворителях, употребляемых для экстракции. Пектины , или пектиновые вещества , при экстракции обычно выделяются первыми, поскольку их растворимость выше. Это - смешанная группа кислых полисахаридов (построенных из моносахаридов арабинозы и галактозы, галактуроновой кислоты, принадлежащей к классу сахарных кислот, и метанола). Длинные молекулы пектиновых веществ могут быть линейными или разветвленными. Срединная пластинка , скрепляющая стенки соседних клеток, состоит из клейких студнеобразных пектатов магния и кальция. В клеточных стенках некоторых созревающих плодов нерастворимые пектиновые вещества превращаются снова в растворимые пектины. При добавлении сахара эти последние образуют гели; поэтому их используют как желирующие вещества.
Гемицеллюлозы - это смешанная группа полисахаридов, растворимых в щелочах (к ним относятся полимеры ксилозы, галактозы, маннозы, глюкозы и глюкоманнозы). У гемицеллюлоз, как и у целлюлозы, молекулы имеют форму цепи, однако их цепи короче, менее упорядочены и сильнее разветвлены.
Клеточные стенки гидратированы: 60-70% их массы обычно составляет вода. По свободному пространству клеточной стенки вода перемещается беспрепятственно. Присутствие воды оказывает влияние на химические и физические свойства полисахаридов клеточной стенки.
Материалы с повышенной механической прочностью, подобные материалу клеточной стенки, т. е. состоящие более чем из одного компонента, называются композиционными материалами или композитами ; их прочность обычно выше, чем у каждого из компонентов в отдельности. Системы из волокон и матрицы (в технике основу композиционного материала называют не матриксом, а матрицей. - Прим. перев) находят широкое применение в технике, так что на изучение их свойств как в технике, так и в биологии тратится много усилий. Матрица, работающая на сжатие, передает напряжение волокнам, работающим на растяжение. Она же обеспечивает абразивную стойкость и, по-видимому, стойкость к неблагоприятным химическим воздействиям, возможным в тех или иных условиях. В строительном деле издавна применяется железобетон, т. е. сочетание бетона со стальной арматурой. Позже появился более легкий композиционный материал, в котором роль матрицы играет пластик, а роль арматуры - стеклянное или углеродное волокно. Древесина представляет собой композиционный материал; своей прочностью она обязана клеточным стенкам. Примером жестких композиционных материалов биологического происхождения могут также служить кость, хрящ и покрывающая экзоскелет членистоногих кутикула. Существуют и гибкие композиционные материалы, например соединительная ткань.
У некоторых клеток, например у клеток мезофилла листа, на всем протяжении их жизни имеется только первичная клеточная стенка. Однако у большинства клеток на внутреннюю поверхность первичной клеточной стенки (кнаружи от плазматической мембраны) отлагаются дополнительные слои целлюлозы, т. е. возникает вторичная клеточная стенка. Обычно это происходит после того, как клетка достигнет своего максимального размера, и лишь немногие клетки, например клетки колленхимы, продолжают рост во время этой фазы. Вторичное утолщение клеточных стенок растения не следует путать с вторичным утолщением (вторичным ростом) самого растения, т. е. с увеличением толщины ствола в результате добавления новых клеток.
В любом слое вторичного утолщения целлюлозные волокна располагаются под одним и тем же углом, но в разных слоях этот угол различен, чем и обеспечивается еще большая прочность структуры. Такое расположение целлюлозных волокон показано на рис. 7.27.
Некоторые клетки, такие, как трахеальные элементы ксилемы и клетки склеренхимы, претерпевают интенсивную лигнификацию (одревеснение); при этом все слои целлюлозы (первичный и три вторичных) пропитываются лигнином - сложным полимерным веществом, не относящимся к полисахаридам. В клетках протоксилемы отложения лигнина имеют кольцевую, спиральную или сетчатую форму, как это видно на рис. 8.11. В других случаях лигнификация бывает сплошной, если не считать так называемых поровых полей, т. е. тех участков в первичной клеточной стенке, через которые осуществляются контакт между соседними клетками при помощи группы плазмодесм (разд. 8.1.3 и рис. 8.7). Лигнин скрепляет целлюлозные волокна и удерживает их на месте. Он действует как очень твердый и жесткий матрикс, усиливающий прочность клеточных стенок на растяжение и в особенности на сжатие (предотвращает прогибы). Он же обеспечивает клеткам дополнительную защиту от неблагоприятных физических и химических воздействий. Вместе с целлюлозой, остающейся в клеточных стенках, лигнин придает древесине те особые свойства, которые делают ее незаменимым строительным материалом.
Функции клеточной стенки
Ниже перечисляются основные функции клеточных стенок растений.
1. Клеточные стенки обеспечивают отдельным клеткам и растению в целом механическую прочность и опору. В некоторых тканях прочность усиливается благодаря интенсивной лигнификации клеточных стенок (небольшое количество лигнина присутствует во всех клеточных стенках).
2. Относительная жесткость клеточных стенок и сопротивление растяжению обусловливают тургесцентность клеток, когда в них осмотическим путем поступает вода. Это усиливает опорную функцию во всех растениях и служит единственным источником опоры для травянистых растений и для таких органов, как листья, т. е. там, где отсутствует вторичный рост. Клеточные стенки также предохраняют клетки от разрыва в гипотонической среде.
3. Ориентация целлюлозных микрофибрилл ограничивает и в известной мере регулирует как рост, так и форму клеток, поскольку от расположения этих микрофибрилл зависит способность клеток к растяжению. Если, например, микрофибриллы располагаются поперек клетки, окружая ее как бы обручами, то клетка, в которую путем осмоса поступает вода, будет растягиваться в продольном направлении.
4. Система связанных друг с другом клеточных стенок (апопласт ) служит главным путем, по которому передвигаются вода и минеральные вещества. Клеточные стенки скреплены между собой с помощью срединных пластинок. В стенках имеются небольшие поры, сквозь которые проходят цитоплазматические тяжи, называемые плазмодесмами . Плазмодесмы связывают живое содержимое отдельных клеток - объединяют все протопласты в единую систему, в так называемый симпласт .
5. Наружные клеточные стенки эпидермальных клеток покрываются особой пленкой-кутикулой, состоящей из воскообразного вещества кутина, что снижает потери воды и уменьшает риск проникновения в растение болезнетворных организмов. В пробковой ткани клеточные стенки по завершении вторичного роста пропитываются суберином, выполняющим сходную функцию.
6. Клеточные стенки сосудов ксилемы, трахеид и ситовидных трубок (с ситовидными пластинками) приспособлены для дальнего транспорта веществ по растению. Этот вопрос рассматривается в гл. 8 и 14.
7. Стенки клеток эндодермы корня пропитаны суберином и поэтому служат барьером на пути движения воды (разд. 14.1.5).
8. У некоторых клеток их видоизмененные стенки хранят запасы питательных веществ; таким способом, например, запасаются гемицеллюлозы в некоторых семенах.
9. У передаточных клеток площадь поверхности клеточных стенок увеличена и соответственно увеличена площадь поверхности плазматической мембраны, что повышает эффективность переноса веществ путем активного транспорта (разд. 14.8.6).
Статьи по теме